Mimicry
In mimicry a message (feature or signal) of one organism, the mimic, resembles some message of another organism which usually belongs to a different species, some feature of the environment, or a generalization of either of those, that is called the model. This resemblance should have some functionality for the mimic by being deceptive for a third participant, the receiver, whose recognition and response is relevant for the mimic. Some researchers use the notion mimicry system to emphasize the systemic nature of mimicry and the relatedness of the three participants.
There is a remarkable variety among mimicry cases in nature. Mimicry exists in most animal classes and also in many plants. Mimetic messages can be transferred in visual, auditory, chemical, tactile and other channels or frequencies that animals use for communication. Mimicry can also be based on many ecological relations such as predation, symbiosis, parasitism, and it can employ different life functions such as foraging, reproduction, and defense.
To organize such high diversity, researchers have proposed many mimicry types and typologies. Historically, the oldest and best-known mimicry types are Batesian mimicry (resemblance of harmless species to some non-edible species that signals their unsuitability to possible predators), Müllerian mimicry (resemblance of aposematic signals of different non-edible species), and aggressive or Peckhamian mimicry (resemblance of predators’ messages to messages of some species, or to some objects, that are harmless to their prey).
Biologists, who have carried out most of the research in mimicry, pay much attention to evolutionary aspects of the phenomenon. The main aspects in the mimicry research of modern biology include: dynamics of mimic and model populations in various selective situations and environmental conditions; behavior of signal receivers with respect to mimics and models, receivers’ abilities to discriminate and learn; variability of mimetic features including genetic and geographical variability of mimics and models. There are also alternative explanations to mimicry that do not rely on evolutionary concepts. Researchers have explained mimicry, for instance, as coincidence because of limited structural combinations in living organisms or as similarity caused by influences of physical conditions in similar living environment.
Also paradigms outside of the natural sciences use the concept of mimicry. For instance in postcolonial studies mimicry has been understood as disruptive imitations that are characteristic of postcolonial cultures. In psychology many authors use mimicry to indicate unconscious imitations between humans, especially related to facial gestures and body movements. In a semiotic context several authors (Sebeok, Nöth, Deacon) have discussed biological mimicry in terms of sign categories and sign processes. There are indeed many directions in mimicry studies, where a semiotic approach can be productive. For instance, semiotics can be applied in analyzing long and complex mimicry displays. Uexküllian biosemiotics, that pays attention to meanings in nature, can be successful in analyzing abstract mimicry, where abstract features, such as ocular shapes, movements or body types common to larger groups, are imitated rather than species-specific characteristics. The Peircean typology of signs opens up new perspectives for classifying mimicry resemblances.
This article was written with the support of Estonian Science Foundation (Grant No 6670).
Introduction
Dictionaries and reference books explain mimicry most often by resemblance; sometimes other clarifying criteria such as concealment, protection or mistaken identity are added. Mimicry can be defined, for instance, as: “the close external resemblance of an animal or plant (or part of one) to another animal, plant, or inanimate object” (NODE 2001: 1175); “the resemblance, through natural selection, of one organism to another or to a natural object, as a natural aid in concealment” (AHD 1981: 834); “the resemblance shown by one animal species, esp. an insect, to another, which protects it from predators” (Collins 1994: 993); “the superficial resemblance that an organism may show to some other animate or inanimate structure, and which serves as a means of concealment” (Trident Webster’s 1995: 635). In general, such descriptions can be considered correct, but like most everyday definitions, they simplify some aspects and leave out others. In the following introduction, I try to describe mimicry not by giving a definition, but by describing neighboring phenomena and thus showing the range of the concept.
First, mimicry or more correctly biological mimicry (to distinguish it from the artistic representations and resemblances in human culture) is considered to be a resemblance between organisms or features of different kinds. That is to say that resemblance between two specimens of the same species (as similarity between human twins), even if it is confusing or used to deceive some third party, does not belong to mimicry. Second, for mimicry to exist, the resemblance should have some functionality for its bearer. There are many accidental resemblances in nature such as the similarity of the chemical compounds of Valerian and Catnip with the sexual attractants for cats. Although confusing and deceptive to cats, such similarity is not mimicry, as there is no functionality involved from the plant’s perspective. Evolutionary biologists often interpret the functionality of mimicry in terms of adaptations and selective advantage of the imitating organism, but one can also describe it in more neutral terms such as the base for different participants to establish permanent relation. Third, singular resemblances such as occasional similarities in gestures or sounds should not be considered mimicry. For mimicry to exist, the resemblance should be repetitive in evolutionary or in an ontogenetic timescale.
On the other hand, mimicry includes many dimensions and phenomena that everyday definitions and understandings usually do not emphasize. For example, the descriptions of mimicry are often limited to the resemblance of colors and forms. Many cases of mimicry are indeed visual. Mimicry studies were launched with the descriptions of visual resemblance between butterfly species (Bates 1862). But mimicry is not limited to visual perception. It can take place in auditory, chemical, tactile or any other channel and frequency that animals use for communication. Related to that, besides being a similarity of body structures, mimicry can also be dynamic and include resemblances of gestures, movements and action patterns. Another widespread understanding is that mimicry is a defense-behavior against predators. Although most mimicry cases are indeed based on predatory relations, mimicry also exists in other ecological relations and functions, such as symbiosis, parasitism or reproduction. The third quite common misunderstanding about mimicry is that the object of imitation should inevitably be a specific species or object. Actually, beside close resemblances with specific biological species (as is common in mimicry between butterflies), more abstract features such as mammalian eyes (eye-spots in many caterpillar and fish) or specific movements (the worm-like movements of an outgrowth of anglerfish) are imitated.
Thus we can specify mimicry not to be a resemblance of one organism to another but rather resemblance of its messages (features or signals) to the messages originating from another being (that usually belongs to different species), to some feature of the environment, or to generalizations of either of those. Such resemblance should be recurrent and deceptive to the third participant in a communicative relationship, the result of which should be relevant to the organism that sends mimetic messages. Unlike most biological definitions of mimicry, the specification given here does not lie on the conceptual ground of evolution theory. Rather it emphasizes that mimicry is a communicative process and thus particularly relevant to semiotics.
Definitions, terminology and uses of the concept
Most research in mimicry has been done in different biological paradigms. This has also shaped the hypotheses raised and questions asked in mimicry studies. Biologists have paid much attention to the evolutionary aspects of the phenomenon — the influence of mimicry resemblance to the participants in the terms of evolutionary success and selection advantage. Evolutionary views have also influenced the definitions used and concepts chosen to explain mimicry.
For instance the British entomologist Richard I. Vane-Wright defines mimicry as follows: “Mimicry occurs when an organism or group of organisms (the mimic) simulates signal properties of a second living organism (the model), such that the mimic is able to take some advantage of the regular response of a sensitive signal-receiver (the operator) towards the model, through mistaken identity of the mimic for the model” (Vane-Wright 1976: 50). The three participants of this definition — mimic, model and operator or (signal) receiver — form the obligatory part of most contemporary theoretical accounts to mimicry. To emphasize the relatedness of the participants in mimicry, Wolfgang Wickler (1965, 1968: 239–242) uses the notion mimicry system to indicate “an ecological setup that includes two or more protagonists, performing three roles” (Pasteur 1982: 169). Different authors and directions of study tend to focus on some particular participant and emphasize its position. For instance, classical ethology and animal psychology prefer to focus on the signal receiver and study its abilities to distinguish similar objects. In early mimicry studies (especially in natural theology, Blaisdell 1982) similarities found by human observers were also used to specify mimicry resemblances; later the position of the animal signal receiver and deception of it has become more prominent as a criterion.
Derbert Wiens, a specialist of plant mimicry, emphasizes the evolutionary approach even more by introducing the Neo-Darwinian concept of fitness to mimicry definition: “I define mimicry as the process whereby the sensory systems of one animal (operator) are unable to discriminate consistently a second organism or parts thereof (mimic) from either another organism or the physical environment (the models), thereby increasing the fitness of the mimic” (Wiens 1978: 367). Andrew Starrett develops this view further by introducing gained fitness as the bases for conceptual unity of mimicry, crypsis, social imitation and other types of resemblances (Starrett 1993). Most extreme Neo-Darwinian approaches explain mimicry in terms of manipulation or exploitation: “natural selection has selected for a resemblance between members of one population A and a second population B, thereby allowing one population to exploit the other” (Hauser 1998: 271).
Systematic mimicry studies were launched together with Darwinian evolution theory in the1860’s, when Henry W. Bates and Alfred R. Wallace used mimicry as a supporting argument for the new evolutionary view of nature (Bates 1862, Wallace 1871). Since that time mimicry studies have been intertwined with the development of evolution theory and interest toward mimicry has followed the ups and downs of Darwinian thought (see Kimler 1986). Besides being a theoretical issue, mimicry has been, however, an important object for empirical studies in modern biology. The main directions in mimicry studies include: dynamics of mimic and model populations in various selection situations and environmental conditions studied both in nature and in computer modeling; behavior of predators and other signal receivers with regard to mimics and models, receivers’ abilities to discriminate and learn difference between mimics and models; variability of mimicry features of species including genetic and geographical variability of mimics and models and so-called mimicry rings (see Bowler 1988, Ruxton, Speed at al. 2004).
Although most biological studies focus on the evolution of mimicry and limit themselves to physical similarity between organisms, there are also definitions, which emphasize communicative and semiotic aspects. The German zoologist Wolfgang Wickler provides a rather semiotically oriented description in the entry in the Encyclopædia Britannica: “Mimicry is a biologic phenomenon characterized by the superficial resemblance of two or more organisms that are not closely related taxonomically. This resemblance confers an advantage — such as protection from predation — upon one or both organisms through some form of ‘information flow’ that passes between the organisms and the animate agent of selection” (Wickler 1998: 144). Also Richard Vane-Wright emphasizes the role of communicative process in mimicry: “[…] the information flow concerning the resemblance between the model and the mimic mediated via the signal-receiver […] is the agent which brings about the mimetic resemblance, or at least maintains or improves it” (Vane-Wright 1976: 28–30).
There are also alternative explanations for mimicry resemblances, which do not rely on natural selection, fitness or other Darwinian concepts (see Komárek 2003). An old tradition rising from German biological philosophy explains mimicry as a coincidence because of the limited number of structural combinations in living organisms (Eimer 1897). Another quite common approach explains mimicry resemblances as the result of the influence of the physical conditions of species that live in similar environments (e.g., Stephenson 1946). Evolutionary developmental biology seems to have given some empirical support to comprehensions that mimicry resemblances may also have non-heritable causes (as in the influence of environmental conditions on early ontogenetic development of butterfly wings, French 1997, Naisbit et al. 2003, Nijhout 1984). There have also been opinions that the functionality of mimicry resemblances is simply human over-interpretation (Heikertinger 1954).
The concept of mimicry is also adapted into cultural studies, partly through biology and partly from the ancient aesthetic concept of mimesis (Aristotle’s Poetics). The Greek word mimēsis is also the etymological origin for the mimicry concept in biology. In postcolonial discourse anthropologist Graham Huggan has described mimicry as aggressive and disruptive imitation, which has the purpose of interfering, ridiculing or subordinating the subject under imitation (Huggan 1997: 94–95). Another well-known postcolonial writer Homi Bhaba uses the notion of mimicry to describe the situation of postcolonial culture “as almost the same, but not quite” and characterizes it by the designations “ambivalence”, “ironic compromise”, “incompleteness” (Bhaba 1994: 122–127). The concept of mimicry is also often used in human psychology to indicate the deceptive or unconscious imitation between humans, especially with reference to facial gestures and body movements (e.g., Gambetta 2005; van Baaren et al. 2004). Such imitation of gestures and movements is important for flexibility of social relations and communication as well as for successful language learning.
In semiotics mimicry is quite probably never explicitly defined, although several authors have used it as an illustrative example or argument. Thomas A. Sebeok has written about mimicry as an example of iconicity in nature: “associations consisting of models and their mimics constitute but one special set of biological events connecting signs in the things signified by ‘a mere relations of reason,’ in which case the sign is an icon” (Sebeok 1989: 116). Following this tradition, other semioticians have related mimicry resemblances with iconicity and iconic signs. For instance Winfried Nöth writes: “visual and olfactory icons occur in the form of mimicry” (Nöth 1990: 124). Terrence W. Deacon considers the relationship between the moth and the tree bark it resembles to be iconic (Deacon 1997: 75–76). A short overview of the concept of mimicry has been published in “S – European Journal for Semiotic Studies” by the mimicry historian Stanislav Komárek (Komárek 1992). In biosemiotics, mimicry as a specific phenomenon has been discussed in relation to recognition and species concept (Kull 1992), intentionality in evolutionary processes (Hoffmeyer 1995), and types of information valuation in communication (Sharov 1992). Proceeding from Peircean semiotics mimicry has been described as a specific type of sign-formation, in which perception and meaning are continually ambivalent to signal receiver (Maran 2005: 125–139).
Diversity, types and typologies
Besides various theoretical explanations and alternate uses of the mimicry concept in different paradigms, mimicry is not a singular or well-delimited phenomenon on the object level. In fact, there is a remarkable diversity among different cases, forms and types of deceptive resemblances in nature and organizing these has been a source of problems and disagreements throughout the history of mimicry studies. There exist many old and well-studied mimicry types such as Batesian, Müllerian and aggressive mimicry that are commonly accepted by the scientific community. Beside those one can also find many novel mimicry types that some individual researchers or schools have used occasionally. Also opinions about the delineation of the mimicry concept and its relations with neighboring phenomena differ. Most authors exclude, for instance, camouflage coloration from mimicry, but the position of biological mimesis, understood as the resemblance with an element of the non-animate environment (stone, twig, leaf, etc.), is more ambivalent. Some authors consider this as a subtype of mimicry whereas others regard it as a separate phenomenon (Endler 1981; Pasteur 1982: 171–173).
Historically, the oldest and best-known mimicry type is Batesian mimicry, named after the British entomologist Henry Walter Bates, who in his fieldwork in the Amazon River described the resemblance of many butterfly species from the families Papilionidae, Pieridae, Lycaenidae a.o. to Ithomia and Methona butterflies (fam. Nymphalidae) (Bates 1862). Batesian mimicry is the resemblance of an edible and harmless species to some poisonous or otherwise non-edible species that signals its unsuitability to possible predators by aposematic coloring or by other conspicuous signals. Besides tropical butterflies Batesian mimicry is well documented not only on other insects such as hover-flies (Howarth, Edmunds 2000; Waldbauer 1988) and ants (Ito, Hashim et al. 2004), but also on snakes (Wüster, Allum et al. 2004), fish (Randall 2005: 301–310), and even on plants (Augner, Bernays 1998).
A contemporary of Bates, the German entomologist Fritz Müller (1878), gave a different explanation of deceptive resemblances. He showed mathematically that it is also advantageous for several unpalatable species to share the same aposematic coloration because the predator then learns more quickly and this, in turn, strengthens the effect of this specific color pattern. This phenomenon has become known as Müllerian mimicry. Some authors argue that this is not actually mimicry and have suggested the term Müllerian convergence, because it is not possible to indicate which species is the mimic and which is the model and who resembles whom (Wickler 1965; Pasteur 1982: 193–194). Also species with an edibility spectrum from palatable to highly noxious may share a similar appearance. Such groups are known as Batesian-Müllerian spectrum or quasi-Batesian mimicry (Huheey 1976; Mallet, Joron 1999).
In aggressive mimicry (also known as Peckhamian mimicry) the mimic does not belong to prey animals but is the predator. Aggressive mimicry helps a predator organism to approach its prey or to lure the prey to approach the predator. Such adaptations are common among spiders (Pekár, Křál 2002), fireflies (Lloyd 1986), some fish (Even, Geoffrey 2004) and carnivorous plants (Moran 1996). The term reproductive mimicry covers various examples, where resemblance aids the mimic in reproduction. Reproductive mimicry is common in many orchids, for instance in fly orchids Ophrys (Schiestl 2005: 257–258). Blossoms of these plants resemble insects by form and smell (Ayasse, Schiestl et al. 2003). They deceive male insects into copulating with the blossoms thereby transferring pollen during this pseudo-copulation. Reproductive mimicry appears also in brood parasitism, for instance in the resemblance of cuckoo eggs to the eggs of their passerine host species (Aviles, Møller 2004; Payne 1977: 8–10). Intraspecific mimicry — understood as mimicry system occurring within one species — also often has a reproductive function. A good example of intraspecific mimicry is the resemblance of the anal fin pattern of the male African mouth-brooding fish Haplochromis burtoni with the eggs of the same species. The resemblance has an important role in spawning: together with the eggs, the female fish also tries to gather the pattern on the male fin into its mouth, and thus catches milt for fertilization (Wickler 1968: 222–227). Within intraspecific mimicry some authors distinguish sexual mimicry as the resemblance between members of different sexes that has functionality in intraspecific communication (Muller, Wrangham 2002).
Besides these categories, there are many mimicry types, which some researchers have used occasionally or which are introduced to denote a specific concept or hypothesis. Behavioral mimicry describes the resemblance of the dynamic behavior of mimic to model (McIver, Stonedahl 1993); instances of behavioral mimicry are locomotor mimicry and escape mimicry (Srygley 1999).Satyric or imperfect mimicry is a partial or approximate resemblance or combination of similar and dissimilar features (Howse, Allen 1994). This mimicry type is believed to operate by causing confusion in signal receivers. Mertensian or Emsleyan mimicry is the name for mimicry complex, where deadly poisonous coral snakes Micrurus imitate moderately poisonous but more numerous species. Such a reverse mimicry system is believed to exist because the learning process is quicker in cases where the receiver remains alive after the encounter (Wickler 1968: 111–121).
A very ambivalent category is floral mimicry or plant mimicry, which unites mimicry cases where the mimic is a plant (Roy, Widmer 1999; Dafni 1984). Actually plants take part in many of the above-mentioned mimicry types such as Batesian, aggressive or reproductive mimicry, but there also exist mimicry types that are specific to plants (see Wiens 1978: 369, 371). In dispersal mimicry the propagules of a plant resemble fruits or other food sources to signal receivers that are usually birds. In weed mimicry weeds or secondary crops resemble crops (flax, wheat), with which they grow. In weed mimicry humans and, in modern times, agricultural machinery serve as signal receivers.
To organize such a high diversity of mimicry, researchers have proposed various typologies. The more common bases for such typologies are: the function of mimicry resemblance to mimic; the nature of the signal or feature being imitated; advantage/disadvantage pattern of resemblance to the participants; species combination involved in mimicry. Many mimicry typologies proceed from Wolfgang Wickler’s concept of the tripartite mimicry system and describe its possible variants. For instance, Richard Vane-Wright (1976: 32–34) and Georges Pasteur (1982: 178) use species combination as a starting point in their mimicry typologies. They distinguish between disjunct mimicry systems where three different species take part, conjunct mimicry systems, where all participants belong to the same species (interspecific mimicry) and three types of bipolar mimicry systems that include two species. In bipolar mimicry systems model and mimic, mimic and receiver or model and receiver can belong to the same species. The latter case occurs for instance in many myrmecomorphic insects that live in ant nests, resemble ants and communicate with ants, who thus also act as the signal receivers (McIver, Stonedahl 1993; Kleisner, Markoš 2005). Mimicry typologies can be quite elaborate. For instance, Vane-Wright’s typology allows distinguishing 40 different mimicry types (Vane-Wright 1976: 34). At the same time classifying mimicry is rarely free of typological problems such as exclusion of some examples of real nature, and dividing others to more than one category (for criticism see Zabka, Tembrock 1986).
Communicational and semiotic aspects
Semiotic studies of biological mimicry have not yet been fully developed and therefore in this overview only some perspectives can be described. To recognize mimicry as a semiotic phenomenon means to focus on perceptual and communicative aspects of mimicry resemblance. In a tripartite mimicry system of mimic, model and signal receiver there are two primary communicative relations (Figure 1). Messages are transmitted from the model to the signal receiver who perceives and interprets these. The mimic releases similar messages. Because of the perceptual similarity of messages from two different sources, the signal receiver has difficulty in distinguishing these. As such a communicational system, mimicry needs to be studied in the framework of other communicational phenomena of nature, such as aposematic colors, species-specific recognition marks and sexual signals; but also by taking into consideration properties of communicational environments, perceptual capabilities of the participating species and ecological relations between these. A suitable paradigm for the semiotic research of biological mimicry is the discipline of zoosemiotics, which investigates semiotic and communicative processes in nature (Sebeok 1972; 1990).
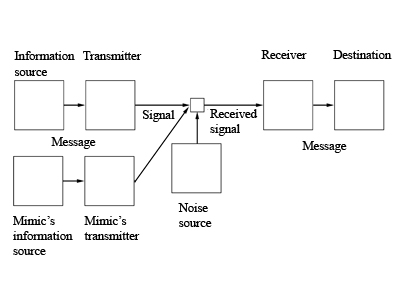
Figure 1. Mimicry system integrated to Shannon-Weaver communication model (modified after Kopp, Mills 2002).
There are several directions in mimicry studies, where a semiotic approach can be productive compared to traditional biological methods. In many mimicry cases the messages sent by the mimic make up an extensive and sometimes controversial set that cannot be treated as a single signal or mimicry type but rather need to be analyzed as a complete display. For example, for the mimicry of anglerfish to be successful, first the inconspicuously colored fish body with fringes and outgrowths needs to match the surrounding environment. In contrast with this a long worm-like appendage wriggles in front of the fish’s mouth. But it is only the combination of cryptic and conspicuous signals, stillness and quick dynamic dash, that makes the aggressive mimicry operational. Such complex mimicry displays can be structured by common dimensions of semiosis — syntactic (focusing on forms), semantic (focusing on meanings) and pragmatic (focusing on use).
A foundation of zoosemiotics is Jakob von Uexküll’s Theory of Meaning. In his writings Uexküll claims that organisms live in their species-specific Umwelten or subjective worlds, and are related with their environment by functional cycles of perception and action, which are organized and mediated by meanings (Uexküll 1982: 27–33). Uexküllian biosemiotics is also relevant to mimicry studies as it allows one to focus on an organism’s perspective in deceptive resemblances instead of describing mimicry as an abstract combination of species. Such a viewpoint helps for instance to understand the phenomenon of abstract mimicry, which has been problematic for biological theory (Pasteur 1982: 191). In abstract mimicry it is not a particular species which is imitated, but a feature common to many species, such as the general form of the eye, a body type or sudden movements (Figure 2). To understand abstract mimicry one should disregard the position that the model is a specific biological species. In abstract resemblance the model for imitation is rather a perception of threat, prey or some other functional category in the receiver’s Umwelt. That perception serves for the signal receiver as the bases of interpretation and through that as the criteria for selection (Maran 2001: 332–334; 2005: 125–139; 2007). In ethology the concept of search image is employed as the set of the object’s representative features that an animal uses in searching for food, mate or some environmental resource. Such search images can also function as a model for resemblance. In semiotic interpretations of mimicry it is necessary to make a distinction between model as the perceptual feature and model species that can embody that feature or transmit the corresponding message.

Figure 2. Abstract mimicry of parasitic flatworm Leucochloridium paradoxum, whose sporocyste tube has invaded the tentacles of the amber snail Succinea. The parasite immobilizes the snail and causes tentacles to pulsate intensely. Brightly colored sporocyste tubes inside the tentacles attract the attention of birds searching for food. This mimicry case belongs to abstract mimicry as it is it is not to possible determine what species is the object of imitation (Photo by Timo Maran, 2002).
In biological discourse there have been lively discussions on the difference between mimicry and cryptic resemblances (see Vane-Wright 1980; Cloudsley-Thompson 1981; Robinson 1981; Endler 1981). This distinction seems to have a semiotic nature and can therefore be efficiently explained using Uexküllian biosemiotics. Namely, the Umwelt of every animal consists of various meaning categories. Some of these correspond to essential functional cycles of sex, food, enemies and shelter. It is vitally important for an animal to apply these, and corresponding cues and signals are actively monitored from the surrounding environment. There is also a neutral background or environment in every Umwelt, which is occasionally noted by the organism but generally ignored during perception. From the semiotic viewpoint the distinction between mimicry and crypsis depends on the position of the model in the receiver’s Umwelt. If the model is important and easily distinguishable for the signal receiver (aposematic coloration, features that correspond to danger, common food items), then it is mimicry. But if the model is the background or informational noise in the receiver’s Umwelt (colors, sounds or smells of the passive environment), then it is crypsis. It seems, that the distinction between mimicry and cryptic resemblance is at least partly relative, depending on the signal receiver’s perception, and therefore it is not possible to make a conclusive distinction from the position of an external observer.
The nature of the model and its relation to the mimic also allows distinguishing mimicry types in a manner similar to the Peircean categories of iconicity, indexicality and symbolicity. As indicated above, it is more common for semiotics to treat mimicry as a sample case of iconicity and iconic signs in nature because of the resemblance of mimic and model as the operating principle of mimicry. The following typology, however, distinguishes different mimicry types according to Peircean categories (Maran 2005: 125–139). Camouflage and crypsis belong to firstness and iconicity, because there is no specific category present in the signal receiver’s Umwelt and the challenge for the receiver is to perceive the imitating organism (this concurs with the Terrence W. Deacon’s opinion that camouflage is essentially iconic as there is no distinction made, Deacon 1997: 75). Classical examples of Batesian mimicry with species-to-species resemblance belong to secondness and indexicality, as there is a distinct category present in the signal receiver’s Umwelt that corresponds to the model and the challenge for the receiver is to determine the type of the found organism. Abstract mimicry belongs to thirdness and symbolicity because the model is some abstract meaning complex in the signal receiver’s Umwelt and the perceivable features related to it. The challenge for the receiver in abstract mimicry is to determine the relation between an encountered organism and its own categories of meaning. By being dependent on the nature of semiosis, the distinction between these types remains relative.
Bibliography
AHD 1981 = Morris, William, ed. (1981). The American Heritage Dictionary of the English Language. Boston: Houghton Mifflin.
Aristotle Poetics = Aristotle. Poetics. Butcher, Samuel H., trans. Mineola: Dover, 1997.
Augner, Magnus; Bernays, Elizabeth A. (1998). “Plant defence signals and Batesian mimicry”. Evolutionary Ecology 12(6): 667–679.
Aviles, Jesús M.; Møller, Anders (2004). “How is host egg mimicry maintained in the cuckoo (Cuculus canorus)?” Biological Journal of the Linnean Society 82(1): 57–68.
Ayasse, Manfred; Schiestl, Florian P. et al. (2003). “Pollinator attraction in a sexually deceptive orchid by means of unconventional chemicals”. Proceedings of the Royal Society of London. Series B–Biological Sciences 270(1514): 517–522.
Bates, Henry W. (1862). “Contributions to an insect fauna of the Amazon valley. Lepidoptera: Heliconidæ”. Transactions of the Linnean Society. Zoology 23: 495–566.
Bhaba, Homi K. (2004). The Location of Culture. London, New York: Routledge.
Blaisdell, Muriel (1982). “Natural theology and Nature’s disguises”. Journal of the History of Biology 15(2): 163–189.
Brower, Lincoln P., ed. (1988). Mimicry and the Evolutionary Process. Chicago: The University of Chicago Press.
Collins 1994 = Collins English Dictionary. 3-th ed. Aylesbury: HarperCollins, 1994.
Cloudsley-Thompson, J. L. (1981). “Comments on the nature of deception”. Biological Journal of the Linnean Society 16: 11–14.
Dafni, A. (1984). “Mimicry and deception in pollination”. Annual Review of Ecology and Systematics 15: 259–278.
Deacon Terrence W. (1997). The Symbolic Species. The Co-Evolution of Language and Brain. New York, London: W. W. Norton.
Eimer, G. M. Theodor (1897). Die Entstehung der Arten auf Grund von Vererben erworbener Eigenschaften nach den Gesetzen organischen Wachsens. Teil 2: Orthogenesis der Schmetterlinge. Jena: G. Fischer.
Endler, John A. (1981). “An overview of the relationships between mimicry and crypsis”. Biological Journal of the Linnean Society 16: 25–31.
Even, Moland; Geoffrey, P. Jones (2004). “Experimental confirmation of aggressive mimicry by a coral reef fish”. Oecologia 140(4): 676–683.
French, Vernon (1997). “Pattern formation in colour on butterfly wings”. Current Opinion in Genetics and Development 7(4): 524–529.
Gambetta, Diego (2005). “Deceptive mimicry in humans”. In: Hurley, Susan; Chater, Nick eds., Perspectives on Imitation. From Neuroscience to Social Science 2: Imitation, Human Development, and Culture. Cambridge: The MIT Press, A Bradford Book, 221–242.
Hauser, Marc D. (1998). The Evolution of Communication. Cambridge: A Bradford Book, The MIT Press.
Heikertinger, Franz (1954). Das Rätsel der Mimicry und seine Lösung. Eine kritische Darstellung des Werdens, des Wesens und der Widerlegung der Tiertrachthypothesen. Jena: Veb Gustav Fisher Verlag.
Hoffmeyer Jesper (1995). “The semiosic body-mind”. Tasca, Norma, ed., Cruzeiro Semiótico 22/25: 367–383.
Howarth, Brigitte; Edmunds, Malcolm (2000). “The phenology of Syrphidae (Diptera): are they Batesian mimics of Hymenoptera?” Biological Journal of the Linnean Society 71(3): 437–457.
Howse, P. E.; Allen, J. A. (1994). “Satyric mimicry: The evolution of apparent imperfection.” Proceedings of the Royal Society of London B 257: 111–114.
Huggan, Graham (1997). “(Post)colonialism, anthropology, and the magic of mimesis”. Cultural Critique 38: 91–106.
Huheey, James E. (1976). “Studies in warning coloration and mimicry VII. Evolutionary consequences of a Batesian–Müllerian spectrum: A model for Müllerian mimicry”. Evolution 30: 86–93.
Ito, Fuminori; Hashim, Rosli, et al. (2004). “Spectacular Batesian mimicry in ants”. Naturwissenschaften 91(10): 481–484.
Kimler William C. (1986). “Mimicry: views of naturalists and ecologists before the modern synthesis”. In Grene, Marjorie, ed., Dimensions of Darwinism. Themes and Counterthemes in Twentieth-Century Evolutionary Theory. Cambridge, UK: Cambridge University Press, 97–127.
Kleisner, Karel; Markoš, Anton (2005). “Semetic rings: Towards the new concept of mimetic resemblances”. Theory in Biosciences 123(3): 209–222.
Komárek, Stanislav (1992). “Mimikry und verwandte Erscheinungen”. S – European Journal for Semiotic Studies 4(4): 693–697.
Komárek, Stanislav (2003). Mimicry, Aposematism and Related Phenomena. Mimetism in Nature and the History of its Study. Muenchen: Lincom Europa.
Kopp, Carlo; Mills, Bruce (2002). “Information warfare and evolution. Conference paper.” In: Proceedings of the 3rd Australian Information Warfare & Security Conference. Online: www.csse.monash.edu.au/~carlo/archive/PAPERS/_JIW-2002-2-CK-BIM-S.pdf
Kull Kalevi (1992). “Evolution and semiotics”. In Sebeok, Thomas A.; Umiker-Sebeok, Jean and Young, Evan P., eds., Biosemiotics. The Semiotic Web 1991. Berlin: Mouton de Gruyter, 221–233.
Lloyd, James E. (1986). “Firefly communication and deception: Oh what a tangled web.” In Mitchell, Robert W.; Thompson, Nicholas S., eds., Deception. Perspectives on Human and Nonhuman Deceit. New York: State University of New York Press, 113–128.
Mallet, James; Joron, Mathieu (1999). “Evolution of diversity in warning color and mimicry: Polymorphisms, shifting balance, and speciation”. Annual Review of Ecology and Systematics 30: 201–233.
Maran, Timo (2001). “Mimicry: Towards a semiotic understanding of nature”. Sign Systems Studies 29.1: 325–339.
Maran, Timo (2005). Mimikri kui kommunikatsioonisemiootiline fenomen. [Mimicry as a communication semiotic phenomenon]. (=Dissertationes Semioticae Universitatis Tartuensis 7). Tartu: Tartu University Press.
Maran, Timo (2007). “Semiotic interpretations of biological mimicry”. Semiotica, forthcoming.
McIver, James D.; Stonedahl, Gary (1993). “Myrmecomorphy – morphological and behavioral mimicry of ants”. Annual Review of Entomology 38: 351–379.
Moran, Jonathan A. (1996). “Pitcher dimorphism, prey composition and the mechanisms of prey attraction in the pitcher plant Nepenthes rafflesiana in Borneo”. Journal of Ecology 84(4): 515–525.
Müller, Fritz (1878). “Ueber die Vortheile der Mimicry bei Schmetterlingen”. Zoologischer Anzeiger 1: 54–55.
Muller, Martin N.; Wrangham, Richard (2002). “Sexual mimicry in hyenas”. The Quarterly Review of Biology 77(1): 3–16.
Naisbit, Russell E.; Jiggins, Chris D.; Mallet, James (2003). “Mimicry: Developmental genes that contribute to speciation”. Evolution and Development 5(3): 269–280.
Nijhout, H. Frederik (1984). “Colour pattern modification by coldshock in Lepidoptera”. Journal of Embryology and Experimental Morphology 81: 287–305.
NODE 2001 = Pearsal, Jude, ed. 2001. The New Oxford Dictionary of English. Oxford: Oxford University Press.
Nöth, Winfried (1990). Handbook of Semiotics. Bloomington: Indiana University Press.
Pasteur, Georges (1982). “A classificatory review of mimicry systems”. Annual Review of Ecology and Systematics 13: 169–199.
Payne, Robert (1977). “The ecology of brood parasitism in birds”. Annual Review of Ecology and Systematics 8: 1–28.
Pekár, Stano; Křál, Jĭri (2002). “Mimicry complex in two central European zodariid spiders (Araneae: Zodariidae): How Zodarion deceives ants”. Biological Journal of the Linnean Society 75(4): 517–532.
Randall, John E. (2005). “A review of mimicry in marine fishes”. Zoological Studies 44(3): 299–328.
Robinson, Michael H. (1981). “A stick is a stick and not worth eating: On the definition of mimicry”. Biological Journal of the Linnean Society 16: 15–20.
Roy, Bitty A.; Widmer, Alex (1999). “Floral mimicry: A fascinating yet poorly understood phenomenon”. Trends in Plant Science 4(8): 325–330.
Ruxton, Graeme D.; Speed, Michael P.; Sherratt, Thomas N. (2004). Avoiding Attack. The Evolutionary Ecology of Crypsis, Warning Signals and Mimicry. Oxford: Oxford University Press.
Schiestl, Florian P. (2005). “On the success of a swindle: pollination by deception in orchids”. Naturwissenschaften 92(6): 255–264.
Sebeok, Thomas A. (1972). Perspectives in Zoosemiotics. (=Janua Linguarum. Series Minor 122). The Hague: Mouton de Gruyter.
Sebeok, Thomas A. (1989). “Iconicity”. In Sebeok, Thomas A. The Sign & Its Masters. Lanham: University Press of America, 107–127.
Sebeok, Thomas A. (1990). Essays in Zoosemiotics. (= Monograph Series of the TSC 5). Toronto: Toronto Semiotic Circle; Victoria College in the University of Toronto.
Sharov, Alexei A. (1992). “Biosemiotics: A functional-evolutionary approach to the analysis of the sense of information”. In Sebeok, Thomas A.; Umiker-Sebeok, Jean and Young, Evan P., eds., Biosemiotics. The Semiotic Web 1991. Berlin: Mouton de Gruyter, 345–373
Srygley, Robert B. (1999). “Incorporating motion into investigations of mimicry”. Evolutionary Ecology 13(7–8): 691–708.
Starrett, Andrew (1993). “Adaptive resemblance: A unifying concept for mimicry and crypsis”. Biological Journal of the Linnean Society 48(4): 229–317.
Stephenson, E. M. (1946). Animal Camouflage. Harmondsworth: Penguin Books.
Trident Webster’s 1995 = The New International Webster’s Dictionary of the English Language. Naples: Trident Press International, 1995.
Uexküll, Jakob v. (1982). “Theory of meaning”. Semiotica 42(1): 25–82.
van Baaren, R. B.; Horgan, T. G.; Chartrand, T. L.; Dijkmans, M. (2004). “The forest, the trees, and the chameleon: Context dependence and mimicry”. Journal of Personality and Social Psychology 86(3): 453–459.
Vane-Wright, Richard I. (1976). “A unified classification of mimetic resemblances”. Biological Journal of the Linnean Society 8: 25–56.
Vane-Wright, Richard I. (1980). “On the definition of mimicry”. Biological Journal of the Linnean Society 13: 1–6.
Waldbauer, G. P. (1988). “Asyncrony between Batesian mimics and their models” In Brower, Lincoln P., ed., Mimicry and the Evolutionary Process. Chicago: The University of Chicago Press, 103–121.
Wallace, Alfred Russell (1871). Contributions to the Theory of Natural Selection. A Series of Essays. 2nd ed. New York: Macmillan and Co.
Wickler, Wolfgang (1965). “Mimicry and the evolution of animal communication”. Nature 208: 519–521.
Wickler, Wolfgang (1968). Mimicry in Plants and Animals. Martin, R. D., trans. London: George Weidenfeld & Nicolson.
Wickler, Wolfgang (1998). “Mimicry”. In The New Encyclopædia Britannica. 15th ed. Macropædia 24, 144–151.
Wiens, Derbert (1978). “Mimicry in plants”. Evolutionary Biology 11: 364–403.
Wüster, Wolfgang; Allum, Christopher S. E. et al. (2004). “Do aposematism and Batesian mimicry require bright colours? A test, using European viper markings”. Proceedings of the Royal Society – Biological Sciences (Series B) 271(1556): 2495–2499.
Zabka, H.; Tembrock, G. (1986). “Mimicry and crypsis — a behavioural approach to classification”. Behavioural Processes 13: 159–176.