Niche Construction
Niche construction can be minimally defined as the process whereby organisms modify their own (and other organisms’) selective environment to such a degree that it changes the selection pressures acting on present and future generations of said organism.
It has long been known to biologists and anthropologists that many animals extensively modify their immediate surroundings. This trend is carried to its extreme in humans, for whom culture has, in fact, become its niche (Hardesty 1972). However, previous models addressing the evolution of such modification behavior and its reciprocal impact on the ecology and evolution of the modifying organism have failed to appreciate the extent to which they may have shaped the evolutionary process by treating them as a by-product of other evolutionary processes (mutation, drift, selection). The niche construction has recently been put forward as a way of integrating ecology and evolution, and to explain the remarkable fit between organisms and their niche. In contrast to previous approaches it contends that the active modification of critical parameters in the external environment of an organism by the organism itself – niche construction – is a significant, yet neglected evolutionary process (Day et al. 2003; Laland and Brown 2006; Laland and Sterelny 2006; Odling-Smee, Laland and Feldman 1996, 2003).
The niche construction approach has also been dubbed the ‘triple-inheritance model’ because it builds on previous so-called dual-inheritance models of cultural evolution (Boyd and Richerson 1985; Richerson and Boyd 2005), which also cast genetic and cultural evolution as by and large independent evolutionary systems. The niche construction approach extends such models by incorporating the ecological domain rather than only addressing cultural traits learned by individuals. The key to the niche construction approach therefore is the inclusion of organism-induced environmental modification bequeathed from the modifying generation to its off-spring. Such a view stands in contrast to traditional, asymmetric definitions of adaptation (e.g., Williams 1992) but allows for two-way adjustments: Either selection leads to changes over time in organisms, or organisms induce changes in their environment in order to off-set further selection (Day et al. 2003; Laland and Brown 2006). Niche construction is rife in the biological world (Odling-Smee et al. 2003) and has profound implications for human bio-social evolution (Laland, Odling-Smee and Feldman 2000, 2001; Odling-Smee 1996, 2006).
Introduction
Niche construction can be minimally defined as the process whereby organism modify their own (and other organisms’) selective environment to such a degree that it changes the selection pressures acting on present and future generations of said organism (Figure 1). Niche construction, also termed ‘triple-inheritance theory’ (Day et al. 2003; Laland and Brown 2006; Laland, Odling-Smee and Feldman 1999, 2000, 2001, 2005; Laland and Sterelny 2006; Odling-Smee 1996, 2006; Odling-Smee, Laland and Feldman 1996, 2003, www.nicheconstruction.com), builds on and extends traditional dual-inheritance models of cultural evolution (e.g., Boyd and Richerson 1985; Laland and Brown 2002; Richerson and Boyd 2005), which have already proven to be highly useful for the quantitative study of human culture. The critical departure of the niche construction/triple-inheritance approach is, however, that organisms are seen to play a more active role in shaping their own and other organisms’ environment, in particular those aspects of the environment that hold selective relevance in an evolutionary perspective. It is argued that the niche construction model serves as a productive general model for human evolution because of its additional emphasis on ecological processes.
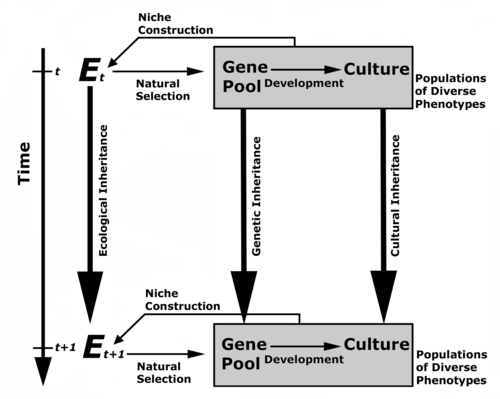
Figure 1. The niche construction or triple-inheritance model of bio-social evolution. In contrast to the standard model of neo-Darwinian evolution, this extended framework also recognizes cultural as well as ecological inheritance as important tiers of human evolution. Cultural inheritance specifically refers to knowledge, skills and artefacts that are in effect personal, whereas ecological inheritance encompasses modification of the ancestral environment that are bequeathed onto the next generation, as well as communally held aspects of culture. Redrawn from Laland et al. (2000: 136).
Genes, Organisms and Environments
The most recent publications on niche construction (Odling-Smee et al. 2003) have been greeted with a somewhat mixed response from biologists (see Keller 2003; Laland, Odling-Smee and Feldman 2004; Okasha 2005; Sterelny 2005; Vandermeer 2004). However, the notion of an interactive relationship between organisms and their environment goes back some time, most pertinently to the writings of Richard Lewontin (1978, 1983, 2000). Adaptation is quite simply defined to encompass “those properties of living things that enable them to survive and reproduce” (Ridley 2004: 6). Although a historical perspective is crucial for understanding adaptations – if something is not shaped through selection it does not qualify as an adaptation – the relationship between the organism and its selecting environment is commonly seen as one-way: “Adaptation is always asymmetrical; organisms adapt to their environments, never vice versa” (Williams 1992: 484). However, in his discussion of the relationship between genes, organisms and environments, Lewontin (1983: 273) notes that…
…what is left out of this adaptive description of organism and environment is the fact, clear to all natural historians, that the environments of organisms are made by the organisms themselves as a consequence of their own life activities.
One can begin to describe niche construction formally as a set of coupled equations. The first part describes the evolutionary change (f) in organisms (O) as a function of the organisms and the environment (E) over time (t; Equation 1).
dO/dt = f(O,E) (1)
The second part describes autonomous change in the state (g) of the environment over time (Equation 2).
dE/dt = g(E) (2)
Whilst there are, of course, autonomous environmental processes that act on, change and disrupt an organism’s niche, the great majority of environmental variables pertinent to selection are local and thus potentially within reach of the selected organism itself. Thus, a closer approximation of the reality of adaptation is a pair of coupled differential equations that combine Equations 1 and 2 and express the effective co-evolution of organisms and their environment:
dO/dt = f(O,E) (3)
dE/dt = g(O,E) (4)
From Equations 3 and 4 it is evident that organism and environment are engaged in a feedback relationship. Whilst organismal evolution is adequately (if somewhat simplistically) expressed by Equation 3, environmental change becomes rather more complex once the active engagement of organisms (O) is taken into consideration. The addition of O to Equation 4 creates a link between environmental change and evolutionary change. Over evolutionary time, such a feedback relationship not only adapts the organism to its environment, but it will also change the environment, so that there is in fact a convergence of the two aspects towards the kind of glove-in-hand fit between living things and their niches (Figure 2). It has also been suggested that such feedback relationships can affect evolutionary dynamics in opposite ways, leading to instability and extinction (Robertson 1991).
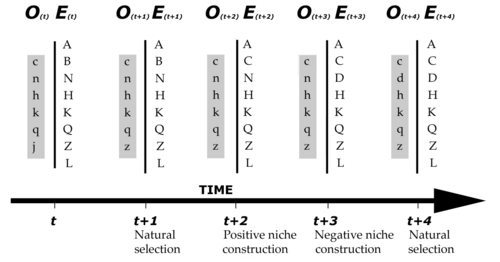
Figure 2. A formal depiction of how both organisms (O) as well as their environment (E) can change over time. The organism is represented by a series of small letters, the environment by capital letter. A match between the letters at each time step (t, t+1, etc.) denotes adaptation, a mismatch lack thereof or mal-adaptation. Redrawn from www.nicheconstruction.com.
Odling-Smee et al. (2003) have amassed a wide range of examples for niche construction in the wild, and more are being accumulated rapidly, ranging from parasites (Boni and Feldman 2005; Patz et al. 2000), to plants (Donohue 2005; Donohue et al. 2005b; Donohue, Polisetty and Wender 2005a; Schwilk 2003), to non-human primates (Byrne et al. 2004) and humans, from both a contemporary (Borenstein, Kendal and Feldman 2006; Ihara and Feldman 2004) as well as an archaeological perspective (Riede 2005a, b). Niche construction occurs at all levels of biotic complexity, but is a particular feature of animals that habitually “construct aspects of their environments, such as nests, holes, burrows, webs, pupal cases” (Day et al. 2003: 81). Beyond these straightforward mechanical manipulations, niche construction activities also often relate to the chemical milieu. However, for the unfolding of the evolutionary potential of this process, organisms “frequently choose, protect, and provision nursery environments for their offspring” (Day et al. 2003: 81) and so immediately affect the often crucial early-stage selective environment of their own descendants.
In order to facilitate the discussion of niche construction processes and the application of the model to case studies, the process itself has been further differentiated. The definitions of the main modes of ecological engineering (synonymous with niche construction) are (Odling-Smee et al. 2003: 418-419; www.nicheconstruction.com):
- Counteractive niche construction involves organisms either perturbing components of their environments, or relocating in their environments in such a way that they wholly or partly reverse or neutralise some prior change in one or more natural selection pressures in their environments. These prior changes in natural selection could be caused either by independent processes in an environment, or by the prior niche constructing activities of other organisms, or by the prior niche-constructing activities of the organisms themselves, and/or their ancestors.
- Inceptive niche construction involves organisms either perturbing components of their environments, or relocating in their environments in such a way that they introduce a new change in one or more natural selection pressures.
- Perturbational niche construction involves organisms physically changing one or more components of their external environments.
- Relocational niche construction involves organisms actively moving in space, as well as choosing or biasing the direction, the distance in space through which they travel, and the time when they travel, thereby modifying natural selection pressures.
These modes of niche construction activities are useful new terms and will be referred to throughout this article. In addition to process definitions, it is also important to recognize niche construction outcomes in terms of their effect on adaptedness. It is particularly pertinent to consider negative niche construction effects. The palaeontological as well as the archaeological record, as O’Brien and Holland (1992) note, are records of evolutionary successes, given that failures are often too short-lived to leave a trace. Yet, ostensibly maladaptive features do exist both in the biological, as well as in the cultural world, and dual-inheritance theory has made considerable headway in explaining the latter. Two points are especially important here: First, behavior that turns out to be adaptively negative can also occur with respect to the ecological parameters of an organism and secondly, the same type of behavior can be negative or positive depending on other variables. Negative niche construction primarily refers to longer term effects of such construction behavior.
- Positive niche construction refers to niche-constructing acts that, on average, increase the fitness of the niche-constructing organisms. In the short run virtually all niche construction by individual organisms is expected to be positive.
- Negative niche construction refers to niche-constructing acts that, on average decrease the fitness of the niche-constructing organisms. In the long run we expect some niche-constructing activities of organisms to become negative for their populations.
Animal Examples of Niche Construction
One of the most instructive examples for the power of niche construction is the case of the common earthworm, Lumbricus terrestris. Its puzzling physiology is described by Turner (2000: 103), who states that “when we apply physiological criteria to deciding what the proper habitat of an earthworm might be, it is hard to escape the conclusion that they are not really terrestrial. Rather, earthworms seem to belong in a fresh-water habitat”. Turner is forced towards this conclusion because earthworm nephridia, the functional equivalent of kidneys in this group of worms, are much closer, both structurally and functionally, to those of fresh-water organisms.
However, earthworms are highly successful organisms, in part because they are powerful mechanical and chemical ecological engineers. They relocate themselves with respect to their environment and have a strong pertubational effect on it. Through the processing of soil and the exuding of mucus, earthworms alter the critical chemical parameters of their environment. In addition, these changes also significantly impact the characteristics of the soil. Thus the purely selfish survival strategy of the earthworm also influences, through its extensive constructionist component, the selective environment for a host of other species (Figure 3). Similar examples from other species abound in nature and are listed encyclopaedically by Odling-Smee et al. (2003). This ubiquity has prompted supporters of the constructionist view to argue for “an overhaul in evolutionary thinking” (Day et al. 2003: 82).
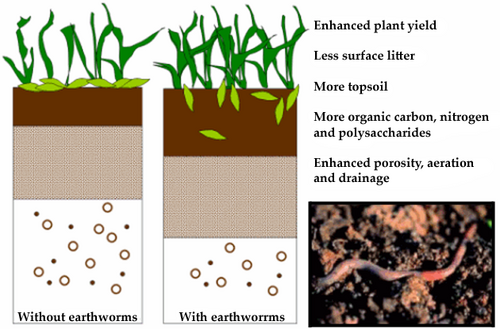
Figure 3. Changes in soil chemistry and composition resulting from the presence and activity of earthworms (far right). The stratigraphic column on the left is a schematic profile through soil without earthworms, the one on the right with earthworms. The humble earthworm powerfully modifies its immediate surroundings and so also changes the selective landscape for a wide variety of other species. Adapted fromwww.nicheconstruction.com.
Niche Construction and the Extended Phenotype
Since the early works of Richard Dawkins (e.g., Dawkins 1976, 1982), it has been widely appreciated that extra-somatic and non-organic components play important roles in the lives and adaptations of many organisms. In order to illustrate how the niche construction view builds on, differs from, and extends Dawkins’ original notions, two further examples of niche construction behaviour are presented: a) spiders’ webs; and b) beavers’ dams. These examples bridge the gap between processes of niche construction observed in the animal kingdom and those observed in human culture.
Spiders and their webs were first discussed at length by Dawkins (1982), who asks his reader to imagine just such a creature sitting at the centre of its web. Although the web is stationary, Dawkins argues, it must be seen as an extension of the spider’s genes, effectively an “extrasomatic means of adaptation” (White 1959: 8). Spiders’ webs are ostensibly adaptive and with the differences in web architecture rooted in spider genes, these differences would also be reflected in genetic differences between populations of spiders that share certain architectural features. In his critical discussion of the notion of the extended phenotype, the anthropologist Anthony Sinclair questions the lead of Dawkins’ gene-centred view of technology and instead argues that one must investigate the social context of production and use in order to understand both human as well as non-human technology. He seems to argue further that technology is primarily “a differential system of value/meaning” (Sinclair 1990: 78), open to political manipulation and largely “conceived as intuitive and implicit” (Sinclair 1990: 78). Sinclair thus advises us to steer clear of evolutionary, selective or adaptive explanations of technology. However, in his initial formulations, Dawkins did not have more than anecdotal evidence about spider web architecture or genetics at his disposable. More recent studies have been able to demonstrate that web genetics and web-building behavior are indeed related (Blackledge and Gillespie 2004). What is more, they can be usefully investigated as behavioral traits within a phylogenetic framework using an explicitly cladistic and quantitative approach (Figure 4). In fact, it is such evolutionary studies of behavioral traits that allow homologous and analogous characters to be discriminated (Benjamin and Zschokke 2004; Wenzel 1992, 1993); scenarios of social and sexual selection can only be reasonably courted only after such convergent adaptations have been taken into consideration, lest we be misled into ‘just so’ interpretations (Gould and Lewontin 1979) or simply be fooled by instances of convergence.
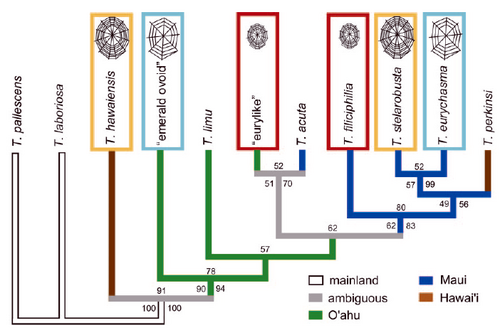
Figure 4. Putting behavioral traits into phylogenetic perspective: Blackledge and Gillespie’s (2004) example of spider web forms on Hawai’i and surrounding islands. The colours denote the various islands and the values associated with each node represent the bootstrap values of the maximum likelihood analysis (left), the maximum parsimony analysis (top) and the posterior probability of the Bayesian approach (right). The coloured boxes indicate web types. From Blackledge and Gillespie (2004: 1229).
Blackledge and Gillespie (2004: 16220-16221) seriously consider what, in the world of spiders, passes for social or contextual interpretations. They state that “sexual selection could play a role in shaping the evolution of tetragnathid web architecture, because courtship and mating take place on the web”. As far as the conclusions of Blackledge and Gillespie and Sinclair go, one could claim that the matter is relatively clear. Human technology is perhaps more complex than spider web architecture, but conceptually the two can be subsumed under the extended phenotype and studied using evolutionary/phylogenetic methods. In fact, the archaeologists O’Brien and Holland (1992, 1995) follow precisely this reasoning, arguing that, with reference to the writings of Dawkins, human-made artefacts are either adaptive or neutral features of the human phenotype.
However, the constructivist argument of Odling-Smee and colleagues complicates the matter somewhat. They argue that the evolutionary significance of spider webs is only incompletely described as an extension of the spider phenotype and thus its genes. Instead, the repeated building of webs has exerted strong selective forces, increasingly changing spider anatomy and other aspects of spider behavior in favour of life on the web. They do, however, also admit that because webs are short-lived, the modification of the spiders’ adaptive landscape would have been primarily through the genetic avenue. The case changes dramatically “when the physical consequences of one generation’s niche construction are not completely erased in the environments of its descendants but are instead bequeathed, either wholly or in part, from one generation to the next, in the form of legacies of modified selection pressures” (Odling-Smee et al. 2003: 13). Such modifications create ecological inheritance and one of the best examples of ecological inheritance, of ecosystem engineering in the wild, is that of beavers and their dams.
Again, beaver dams were introduced into this debate by Dawkins (1976, 1982), and it is worth quoting at length from the Extended Phenotype (Dawkins 1982: 200):
A beaver that lives by a stream quickly exhausts the supply of food trees lying along the stream bank within a reasonable distance. By building a dam across the stream the beaver creates a large shoreline which is available for safe and easy foraging without the beaver having to make long and difficult journeys overland. If this interpretation is right, the lake may be regarded as a huge extended phenotype, extending the foraging range of the beaver in a way which is somewhat analogous to the web of the spider. As in the case of the spider web, nobody has done a genetic study of beaver dams, but we really do not need to in order to convince ourselves of the rightness of regarding the dam, and the lake, as part of the phenotypic expression of beaver genes. It is enough that we accept that beaver dams must have evolved by Darwinian natural selection.
Dawkins’ reasoning is again echoed by O’Brien and Holland (1992, 1995), who would transpose this phenotypic extension onto human artefacts under the “premise that things contained in the [archaeological] record were parts of past human phenotypes” and that “importantly, the objects are not simply reflective of or related to phenotypes…they are parts of phenotypes in the same way beaver dams and bird nests are parts of phenotypes” (O’Brien and Holland 1992: 37, emphasis added). Indeed, they perhaps somewhat prematurely claim that “without this tenet the application of evolutionary theory to the understanding of how things found in the archaeological record came to be the way they are makes absolutely no sense” (O’Brien and Lyman 2000: 141). But in what way exactly is a beaver’s dam part of the beaver phenotype? Dawkins did not fully appreciate the evolutionary significance of beaver dams. These constructs outlive several generations of beavers and thus have a generational length above that of beaver genes. Odling-Smee (1996) and Odling-Smee et al. (2003) argue that this is a prime instance of ecosystem engineering and ecological inheritance, where a new generation of beavers is born into an environment that has been subject to the active manipulation by its ancestors. Furthermore, beaver lodges are critical for the survival of offspring and impact significantly on breeding behavior and success. Beaver genes, beaver phenotype and beaver environment thus are coupled, as in Equations 3 and 4, and cannot be understood fully without reference to all three aspects of beaver existence. In addition, the dam building behavior of beavers also impacts on the selective regimes of other species (e.g., Wright, Jones and Flecker 2002), another key feature of the niche construction model.
Genetic data on beavers are only now becoming available (Milishnikov 2004) and an appreciation of the interconnectedness of the beaver niche with that of other organisms (e.g., Wright et al. 2002), including humans (e.g., Coles 2006), is growing. Unfortunately, nobody has as yet conducted a phylogenetic analysis of beavers using dam architecture as a behavioral trait. However, of great interest to human scientists, beaver dams have been addressed in terms of the so-called chaîne opératoire model of cultural-technological competence (see Bleed 2001; Riede 2006; Schlanger 1994). Karlin and Julien (1994: 152) note that operational sequences of technological production, manipulation and use do not only exist in human culture, but also in nature: “the building of a nest, or a beaver lodge, and the hoarding of food supplies for hibernation”. Such an approach would collapse animal and human technology under the heading of the extended phenotype. But if Odling-Smee (1996) and Odling-Smee et al. (2003) are correct in pointing out the limits of phenotypic extensions, then a strict insistence on this theory by evolutionarily-minded human scientists will inevitably be incomplete too. Sterelny (2006: 151-152) agrees that “to the extent that information does flow collectively, niche construction is our best model of the generation-by-generation accumulation of skill, technology and information”. He goes on to argue that dual-inheritance theory provides a robust theory for examining the flow of information in cases where it is not collectively held. Such a distinction between personal and collective knowledge and material culture mirrors Oswalt’s (1976) categorization of human technology into instruments and weapons on the one hand and tended and untended facilities on the other. Instruments/weapons reflect the social transmission of information along personal lines, whilst facilities reference the collective flow of information in human societies (cultural and ecological inheritance in Figure 1). It is the latter types that have been argued to be linked to the long-term human population growth and density and to play a key role in cultural evolution. Conceptualizing the human built environment and the collective flow of information as part and parcel of on-going niche construction process in human evolution casts new light on the evolution of this critical trait of Homo sapiens.
The general properties of the niche construction, or triple-inheritance model have been outlined, first in general terms and then with regards to non-human plants and animals. On the basis of this discussion, it has been argued that the model is also highly applicable to human cultural evolution, noting that “culture and niche construction have become self-reinforcing, with transgenerational culture modifying the environment in a manner that favours ever more culture, and niche construction informed by cultural knowledge becoming ever more powerful” (Laland and Brown 2006: 96). Mathematical models have indicated that even slight but sustained and inherited modifications of the ancestrally selective environment can have a dramatic impact on evolution in general, and human evolution in particular, but as Vandermeer (2004: 473) notes “including considerations of culture complicates but does not really alter the basic niche construction approach. The gene-culture co-evolution point of view…is thus enriched substantially”. Any evolutionary model demands a diachronic perspective in order to address questions of heritability and adaptation. Archaeology and anthropology are ideally situated to contribute this longue durée to niche construction arguments regarding human cultural evolution, and the niche construction model offers a potentially unifying approach for the study of human bio-social evolution (Mesoudi, Whiten and Laland 2006). Echoing the arguments by Shennan (1989, 2002, 2005), Vanpool and Vanpool (2003) and Kuhn (2004), evolutionary approaches to culture are not presented as rival theories of culture change, but rather as complementary to other forms of inquiry. In a sense, the niche construction approach provides a vocabulary and methodology for a more integrated evolutionary perspective on human evolution. This perspective is commensurable with social and cultural understandings of human society and does not posit causality with the external environment a priori, partly because the notion of an external environment itself is deconstructed. The observation that we cannot sideline environmental change in our discussion of cultural change therefore does not stem from a position of vulgar environmental determinism, but from the empirical observation that large-scale and independent climate change and archaeologically visible changes in human culture are often strongly correlated. Perhaps the primary adaptive effect of human culture is to offset these environmental perturbations (Alvard 2003; Laland and Brown 2006).
Niche construction has hitherto been a ‘neglected process in evolution’, but as Laland et al. (2001: 32) note there are “rich opportunities to explore the impact of cultural niche construction in human evolution” through archaeological, anthropological and psychological evidence. As was noted earlier, many biologists have reservations regarding the niche construction approach. Laland and Brown (2006: 97) acknowledge that “it remains to be seen whether this perspective will become an established feature of evolutionary theory”, yet there are many reasons not to further ignore this approach (see Laland and Sterelny 2006).
Bibliography
Alvard, M.S. (2003). The Adaptive Nature of Culture. Evolutionary Anthropology 12: 136-49.
Benjamin, S.P. and S. Zschokke (2004). Homology, behaviour and spider webs: web construction behaviour of Linyphia hortensis and L. triangularis (Araneae: Linyphiidae) and its evolutionary significance. Journal of Evolutionary Biology 17: 120-30.
Blackledge, T.A. and R.G. Gillespie (2004). Convergent evolution of behavior in an adaptive radiation of Hawaiian web-building spiders. Proceedings of the National Academy of Sciences of the United States of America 101: 16228-23.
Bleed, P. (2001). Trees or Chains, Links or Branches: Conceptual Alternatives for Consideration of Stone Tool Production and Other Sequential Activities. Journal of Archaeological Method and Theory 8: 101-27.
Boni, M.F. and M.W. Feldman (2005). Evolution of antibiotic resistance by human and bacterial niche construction. Evolution 59: 477-91.
Borenstein, E., J. Kendal and M. Feldman (2006). Cultural niche construction in a metapopulation. Theoretical Population Biology 70: 92-104.
Boyd, R. and P.J. Richerson (1985). Culture and the evolutionary process. Chicago, IL.: University of Chicago Press.
Byrne, R.W., P.J. Barnard, I. Davidson, V.M. Janik, W.C. McGrew, A. Miklosi and P. Wiessner (2004). Understanding culture across species. Trends in Cognitive Sciences 8: 341-46.
Coles, B.J. (2006). Beavers in Britain’s Past. Oxford: Oxbow.
Dawkins, R. (1976). The Selfish Gene. Oxford: Oxford University Press.
Dawkins, R. (1982). The Extended Phenotype. The Long Reach of the Gene. Oxford: Oxford University Press.
Day, R.L., K.N. Laland, J. Odling-Smee and M.W. Feldman (2003). Rethinking Adaptation: The Niche Construction Perspective. Perspectives in Biology and Medicine 46: 80-95.
Donohue, K. (2005). Niche construction through phenological plasticity: life history dynamics and ecological consequences. New Phytologist 166: 83-92.
Donohue, K., C.R. Polisetty and N.J. Wender (2005a). Genetic basis and consequences of niche construction: plasticity-induced genetic constraints on the evolution of seed dispersal in Arabidopsis thaliana. American Naturalist 165: 537-50.
Donohue, K., L. Dorn, C. Griffith, E. Kim, A. Aguilera, C.R. Polisetty and J. Schmitt (2005b). Niche construction through germination cueing: life-history responses to timing of germination in Arabidopsis thaliana. Evolution 59: 771-85.
Gould, S.J. and R.C. Lewontin (1979). The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proceeding of the Royal Society of London, Series B 205: 581-98.
Hardesty, D.L. (1972). The Human Ecological Niche. American Anthropologist 74: 458-66.
Ihara, Y.W. and M. Feldman (2004). Cultural niche construction and the evolution of small family size. Theoretical Population Biology 65: 105-11.
Karlin, C. and M. Julien (1994). “Prehistoric technology: a cognitive science?” in Renfrew, C. and E.B.W. Zubrow eds., The ancient mind. Elements of cognitive archaeology. Cambridge: Cambridge University Press.(152-64)
Keller, L. (2003). Changing the world. Nature 425: 769-70.
Kuhn, S.L. (2004). Evolutionary perspectives on technology and technological change. World Archaeology 36: 561-70.
Laland, K.N. and G.R. Brown (2002). Sense & Nonsense. Evolutionary Perspectives on Human Behaviour. Oxford: Oxford University Press.
Laland, K.N. and G.R. Brown (2006). Niche Construction, Human Behavior, and the Adaptive-Lag Hypothesis. Evolutionary Anthropology 15: 95-104.
Laland, K.N. and K. Sterelny (2006). Perspective: Seven Reasons (Not) to Neglect Niche Construction. Evolution 60: 1751-62.
Laland, K.N., F.J. Odling-Smee and M.W. Feldman (1999). Evolutionary consequences of niche construction and their implications for ecology. Proceedings of the National Academy of Sciences of the United States of America 96: 10242-47.
Laland, K.N., F.J. Odling-Smee and M.W. Feldman (2000). Niche construction, biological evolution, and cultural change. Behavioural and Brain Sciences 23: 131-75.
Laland, K.N., F.J. Odling-Smee and M.W. Feldman (2001). Cultural niche construction and human evolution. Journal of Evolutionary Biology 14: 22-33.
Laland, K.N., F.J. Odling-Smee and M.W. Feldman (2004). Causing a commotion. Nature 429: 609.
Laland, K.N., F.J. Odling-Smee and M.W. Feldman (2005). On the Breath and Significance of Niche Construction: A Reply to Griffiths, Okasha and Sterelny. Biology and Philosophy 20: 37-55.
Lewontin, R.C. (1978). Adaptation. Scientific American 239: 156-69.
Lewontin, R.C. (1983). “Gene, organism and environment”. in Bendall, D.S. ed. Evolution from molecules to men. Cambridge: Cambridge University Press.(273-86)
Lewontin, R.C. (2000). The Triple Helix. Gene, Organism and Environment. Cambridge, MA: Harvard University Press.
Mesoudi, A., A. Whiten and K.N. Laland (2006). Towards a unified science of cultural evolution. Behavioural and Brain Sciences 29: 329-83.
Milishnikov, A.N. (2004). Population-Genetic Structure of Beaver ( Castor fiber L., 1758) Communities and Estimation of Effective Reproductive Size N e of an Elementary Population. Russian Journal of Genetics 40: 772-81.
O’Brien, M.J. and T.D. Holland (1992). The Role of Adaptation in Archaeological Explanation. American Antiquity 57: 36-59.
O’Brien, M.J. and T.D. Holland (1995). “Behavioural Archaeology and the Extended Phenotype”. in Skibo, J.M., W.H. Walker and A.E. Nielsen eds., Expanding Archaeology. Salt Lake City, UT.: University of Utah Press.(143-61)
O’Brien, M.J. and R.L. Lyman (2000). “Evolutionary Archaeology. Reconstructing and Explaining Historical Lineages”. in Schiffer, M.B. ed. Social Theory in Archaeology. Salt Lake City, UT.: University of Utah Press.(126-42)
Odling-Smee, F.J. (1996). Niche construction, genetic evolution and cultural change. Behavioural Processes 35: 195-205.
Odling-Smee, F.J. (2006). “How Niche Construction Contributes to Human Gene-Culture Coevolution”. in Wells, J.C.K., S. Strickland and K.N. Laland eds., Social Information Transmission and Human Biology. London: CRC Press.(39-58)
Odling-Smee, F.J., K.N. Laland and M.W. Feldman (1996). Niche Construction. American Naturalist 147: 641-48.
Odling-Smee, F.J., K.N. Laland and M.W. Feldman (2003). Niche Construction. The Neglected Process in Evolution. Princeton: Princeton University Press.
Okasha, S. (2005). On Niche Construction and Extended Evolutionary Theory. Biology and Philosophy 20: 1-10.
Oswalt, W.H. (1976). An Anthropological Analysis of Food-Getting Technology. New York, N.Y.: John Wiley & Sons.
Patz, J.A., T.K. Graczyk, N. Geller and A.Y. Vittor (2000). Effects of environmental change on emerging parasitic diseases. International Journal of Parasitology 30: 1395-405.
Richerson, P.J. and R. Boyd (2005). Not by genes alone: how culture transformed human evolution. Chicago: University of Chicago Press.
Ridley, M. (2004). Evolution. Oxford: Blackwell.
Riede, F. (2005a). “Darwin vs. Bourdieu. Celebrity Deathmatch or Postprocessual Myth? Prolegomenon for the Reconciliation of Agentive-Interpretative and Ecological-Evolutionary Archaeology”. in Cobb, H. et al. eds., Investigating Prehistoric Hunter-Gatherer Identities: Case Studies from Palaeolithic and Mesolithic Europe. Oxford: Oxbow.(45-64)
Riede, F. (2005b). To Boldly Go Where No (Hu-)Man Has Gone Before. Some Thoughts on The Pioneer Colonisations of Pristine Landscapes. Archaeological Review from Cambridge 20: 20-38.
Riede, F. (2006). Chaîne Opèratoire – Chaîne Evolutionaire. Putting Technological Sequences in Evolutionary Context. Archaeological Review from Cambridge 21: 50-75.
Robertson, D.S. (1991). Feedback Theory and Darwinian Evolution. Journal of Theoretical Biology 152: 469-84.
Schlanger, N. (1994). “Mindful technology: unleashing the chaîne opèratoire for an archaeology of mind”. in Renfrew, C. and E.B.W. Zubrow eds., The ancient mind. Elements of cognitive archaeology. Cambridge: Cambridge University Press.(143-51)
Schwilk, D.W. (2003). Flammability is a niche construction trait: canopy architecture affects fire intensity. American Naturalist 162: 725-33.
Shennan, S.J. (1989). “Cultural transmission and cultural change”. in Van Der Leeuw, S. and R. Torrence eds., What’s New? A Closer Look at the Process of Innovation. London: Routledge.(330-46)
Shennan, S.J. (2002). Genes, Memes and Human History: Darwinian Archaeology and Cultural Evolution. London: Thames and Hudson.
Shennan, S.J. (2005). Culture, society and evolutionary theory. Archaeological Dialogues 11: 107-14.
Sinclair, A. (1990). Technology as Phenotype? An Extended Look at Time, Energy and Stone Tools. Archaeological Review from Cambridge 9: 71-81.
Sterelny, K. (2005). Made By Each Other: Organisms and Their Environment. Biology and Philosophy 20: 21-36.
Sterelny, K. (2006). Memes Revisited. The British Journal for the Philosophy of Science 57: 145-65.
Turner, J.S. (2000). The extended organism: the physiology of animal-built structures. Cambridge, MA.: Harvard University Press.
Vandermeer, J. (2004). The Importance of a Constructivist View. Science 303: 472-74.
VanPool, T.L. and C.S. VanPool (2003). “Agency and Evolution: The Role of Intended and Unintended Consequences of Action”. in Vanpool, T.L. and C.S. Vanpool eds., Essential Tensions in Archaeological Method and Theory. Salt Lake City, UT.: University of Utah Press.(89-114)
Wenzel, J.W. (1992). Behavioral homology and phylogeny. Annual Review of Ecology and Systematics 23: 361-81.
Wenzel, J.W. (1993). Application of the biogenetic law to behaviour ontogeny: a test using nest architecture in paper wasps. Journal of Evolutionary Biology 6: 229-47.
White, L.A. (1959). The Evolution of Culture. New York, N.Y.: McGraw-Hill.
Williams, G.C. (1992). Gaia, Nature Worship and Biocentric Fallacies. The Quarterly Review of Biology 67: 479-86.
Wright, J.P., C.G. Jones and A.S. Flecker (2002). An ecosystem engineer, the beaver, increases species richness at the landscape scale. Oecologia 132: 96-101.